INTRODUCCIÓN
En estudios in vivo (1) e in vitro (2-5) se ha observado que el estrés calórico compromete la competencia del ovocito para ser fertilizado (1) y el desarrollo del embrión hasta el estadio de blastocisto (3,4,5). Se ha descrito que la susceptibilidad del embrión al estrés calórico se da en los primeros tres días de edad (6); y que los embriones desarrollados in vitro son más resistentes a este efecto cuando presentan un desarrollo de 4 a 8 células (7,8,9).
La transferencia embrionaria se ha visto como una alternativa para incrementar la tasa de gestación de las vacas lecheras en épocas cálidas. Varios estudios realizados en vacas receptoras de embriones frescos producidos in vitro (10,11) o de embriones recolectados de vacas superovuladas (12,13,14) han informado una mejora de la tasa de gestación en condiciones de estrés calórico, comparada con la observada en vacas servidas con inseminación artificial.
Existen pocas investigaciones diseñadas para determinar un efecto de interacción del estrés calórico de la época de producción del embrión y la época de transferencia sobre la tasa de gestación de vacas receptoras lactantes. En algunos estudios realizados durante el verano no se observó mejora de la tasa de gestación de vacas receptoras de embriones producidos en condiciones de confort térmico(13,15) e incluso se mostró una reducción de este parámetro(16).
Los objetivos del estudio fueron determinar en vacas Holstein lactantes durante su pico de lactancia, el efecto de interacción del estrés calórico de la época de producción del embrión y la de transferencia sobre la tasa de gestación de vacas receptoras, y evaluar si existe efecto del estrés calórico sobre la producción y calidad embrionaria en vacas superovuladas.
MATERIALES Y MÉTODOS
El estudio se realizó en dos explotaciones comerciales de ganado lechero del altiplano central de México (Aguascalientes, México; 22° 05´ N y 102° 16’ O). La estación climática estuvo localizada entre ambos hatos lecheros a una distancia de 5 km, en el municipio de San Francisco de los Romo, Aguascalientes.
La alimentación se proporcionó seis veces al día en forma de ración integral. La relación forraje: concentrado fue de 35:65. La composición de la dieta contenía en base seca: 1.79 Mcal/kg de energía neta de lactancia; 17.7 % de proteína cruda, de la que el 37 % fue no degradable; 19.6 % de fibra detergente ácido; 35 % de fibra detergente neutro; 7.1 % de grasa y 31 % de carbohidratos no estructurales. Las vacas tuvieron libre acceso al agua y a una mezcla de sales minerales.
La superovulación y la colección embrionaria fueron realizadas en dos épocas: 1) Templada, los tratamientos superovulatorios iniciaron el 5 de febrero (n=10) y el 12 de marzo (n=10) y la colección embrionaria se realizó el 17 de febrero y el 24 de marzo, respectivamente; 2) Cálida, los tratamientos superovulatorios iniciaron el 21 de mayo (n=10) y el 2 de julio (n=12), y la colección embrionaria se realizó el 2 de junio y el 14 de julio, respectivamente.
Colección embrionaria
Se utilizaron 42 vacas lactantes Holstein clínicamente sanas de dos hatos comerciales de producción de leche, las cuales se ordeñaron tres veces al día sin recibir durante el período de estudio somatotropina bovina. Al inicio del estudio, de las vacas donadoras se registraron el número de partos, los días en leche la producción de leche estimada a 305 días (P305), la condición en una escala de 1 a 5(17) y el peso corporal en ambas épocas de superovulación y colección embrionaria .
En cada época de colección embrionaria se registró la temperatura ambiente (°C) y la humedad relativa (HR) en una estación climática local. La temperatura registrada en °C se transformó a grados Fahrenheit (°F). Se calculó el índice temperatura–humedad (THI) utilizando la temperatura máxima y la humedad relativa promedio(18), con la siguiente ecuación:
THI = °F – [(0.55 – ((HR/100) x 0.55)) * (°F - 58)]
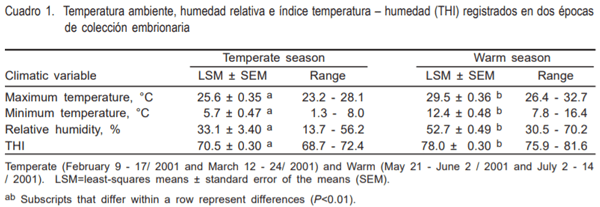
En el Cuadro 1, se observa que los valores medios de temperatura ambiente (29.5 ± 0.4 °C), humedad relativa (52.9 ± 3.5) y THI (78.0 ± 0.3) de la época cálida fueron superiores (P<0.01) a los observados en la templada (25.6 ± 0.3, 33.1 ± 3.4 y 70.5 ± 0.3, respectivamente)
El tratamiento superovulatorio inició entre 9 y 11 días después del estro con un total de 400 mg de hormona folículo estimulante (Folltropin–V, Vetrepharm, Inc.) del mismo lote de producción por vía im cada 12 h durante cuatro días usando dosis decrecientes(19). Al tercer día del tratamiento se administraron por vía im 50 mg prostaglandina F2α (Lutalyse, Pharmacia & Upjohn, Inc.) fraccionadas en dos aplicaciones, una por la mañana y otra por la tarde. La detección del estro se realizó en forma continua con el apoyo de detectores de monta que se activaban a la presión (Kamar Heatmount® Detector, Kamar Inc.). La inseminación artificial se realizó a las 12 y 24 h de detectada la primera monta homosexual.
Siete días después de la inseminación artificial, se realizó un examen genital y se determinó el número de cuerpos lúteos presentes en ambos ovarios. Los embriones fueron recuperados del útero en forma no quirúrgica mediante lavados con una solución salina amortiguada con fosfatos (SSF) y suplementada con 0.4% de albúmina sérica bovina, usando una sonda Foley de dos vías (Unomedical, Bakar Arang Industrial). La colección embrionaria se realizó por un solo profesional calificado. El efluente del lavado de los cuernos uterinos pasó a través de un filtro de 75 µM (Embryo MiniflushTM System). El filtro fue lavado y decantado a una caja de Petri cuadriculada 100 x 100 mM (Falcon Inc.) para la búsqueda de los embriones y óvulos, con la ayuda de un microscopio estereoscópico con 12 aumentos.
Los embriones recolectados se evaluaron conforme a lo descrito en el Manual de Procedimientos de la Sociedad Internacional de Transferencia de Embriones(20), de acuerdo a su estadío de desarrollo y calidad. Se congelaron exclusivamente embriones del estadío de blastocisto de calidad excelente o buena con el crioprotector ethylene glycol (AB Technology Inc.). Los embriones se equilibraron en SSF con 1.5 M de ethylene glycol por un período máximo de 5 min, tiempo en el cual los embriones fueron empajillados. Las pajillas con los embriones fueron colocadas en una congeladora manual previamente enfriada a –6 °C. Después de un minuto se indujo la cristalización; posteriormente la temperatura se llevó hasta -30 ° C a una tasa de -0.5 °C/min, sumergiéndose después las pajillas en nitrógeno líquido.
Los embriones producidos y congelados (n=107) durante la época templada (PT) y cálida (PC); de calidad excelente (n=66) y buena (n=41) fueron distribuidos al azar, dentro del mismo hato a dos épocas de transferencia (T): 1) Templada (TT), del 15 de enero al 27 de marzo; 2) Cálida (TC), del 1 de mayo al 17 de agosto. De esta forma se formaron los siguientes grupos experimentales: producción templada – transferencia templada (PTTT); producción templada – transferencia cálida (PT-TC); producción cálida – transferencia templada (PC-TT); y producción cálida – transferencia cálida (PC-TC).
Transferencia de embriones
Como receptoras se utilizaron 107 vacas Holstein lactantes con tres ordeños al día sin recibir durante el período de estudio somatotropina bovina. El día de la transferencia se registraron de las vacas receptoras el número de partos, los días en leche, la condición y el peso corporal y la P305 en ambas épocas de transferencia.
La temperatura y la humedad del ambiente se registraron en una estación climática local. Los valores medios de temperatura máxima (28.2 ± 0.3), humedad relativa (58.7 ± 1.4) y THI (76.5 ± 0.5) registrados en la época cálida fueron superiores a los observados en la época templada (24.5 ±0.3; 45.5 ± 1.7 y 70.1 ± 0.5, respectivamente) (P<0.01) (Cuadro 2).
La transferencia embrionaria se realizó al séptimo día después del estro. Los embriones se descongelaron a temperatura ambiente por 10 seg seguida de la inmersión en agua a una temperatura de 30 °C por 12 seg(13,21). Los embriones fueron transferidos en forma no quirúrgica y se depositaron en el tercio anterior del cuerno uterino ipsilateral al cuerpo lúteo. En cada evento de transferencia se registró el tiempo transcurrido (min) desde el momento de la descongelación hasta la finalización de la transferencia. Posteriormente, las vacas receptoras se observaron dos veces al día para la detección del estro hasta que se realizó el diagnóstico de gestación por palpación rectal entre 40 a 45 días de edad del embrión.
El día de la transferencia del embrión se obtuvo una muestra sanguínea de las vacas receptoras en un tubo Vacutainer sin heparina (Tyco Healthcare Group LP) y fue transportada al laboratorio en refrigeración a 5 °C. Al día siguiente por la mañana, las muestras se centrifugaron a 700 xg por 10 min, y los sueros fueron almacenados a –20 °C hasta su procesamiento en el laboratorio de radioinmunoensayo para la medición de la concentración sérica de progesterona (Coat-A-Count Progesterone, Diagnostic Products Corporation, Los Angeles, CA, USA). A dosis de 0.49 ng/ml el sistema presentó una sensibilidad de 0.1 ng/ml, con un coeficiente de variación intra e interensayo de 3.6 y 5.8 %, respectivamente. A dosis de 20.0 ng/ml, los coeficientes de variación intra e interensayo fueron de 5.6 y 6.1%, respectivamente.
Variables evaluadas
Las variables evaluadas fueron: porcentaje de vacas que respondieron al tratamiento superovulatorio; número de cuerpos lúteos detectados al momento de la colección embrionaria; tasa de fertilización (número de embriones entre el número de embriones y óvulos); número de embriones más óvulos, número de embriones y número de óvulos colectados por vaca; tasa de colección de embriones más óvulos (número de embriones más óvulos colectados entre el número de cuerpos lúteos); tasa de colección embrionaria (número de embriones entre el número de cuerpos lúteos); distribución porcentual de las vacas superovuladas en relación al número de embriones y óvulos recolectados; grado de desarrollo y calidad de los embriones recolectados; y tasa de gestación de las vacas receptoras de embriones.
Análisis estadístico
Para determinar la similitud de las características productivas de las vacas donadoras y receptoras al inicio del estudio en ambas épocas, como son: el número de partos; los días en leche; la condición y el peso corporal; la producción estimada de leche a 305 días y la concentración sérica de progesterona de la vaca receptora al momento de la transferencia se evaluaron por medio de un análisis varianza con bloques al azar. El modelo al que se atribuyó la varianza consideró al hato como bloque y la época de colección embrionaria y la de transferencia como efecto principal(22).
Las variables climáticas de las épocas de colección embrionaria y transferencia se analizaron por análisis de varianza, donde la época de colección embrionaria o de transferencia se consideró como efecto principal(22).
La respuesta superovulatoria; número de cuerpos lúteos; tasa de fertilización; número de embriones y número de óvulos; tasa de colección de embriones más óvulos; tasa de colección de embriones; distribución porcentual de los embriones por grado de desarrollo y calidad; y la distribución porcentual de las vacas superovuladas en relación al número de embriones y óvulos recolectados se analizaron con Ji-cuadrada(22).
El valor esperado de la tasa de gestación de las vacas receptoras (Y) se analizó con un modelo de regresión logística múltiple de primer orden y representado como:
E{Y} = [1 + exp (-β’ X)]-1
Donde:
β’X-1 = β0 + β1(P)1 + β2(T)2 + β12(P x T) + β3(EQ)3 + β4(P305)4 + β5(ETT)5
Este modelo fue ajustado por el método de máxima verosimilitud; donde los efectos considerados fueron: época de producción del embrión (P), época de transferencia del embrión (T), interacción de las épocas de producción y de transferencia (P x T), calidad del embrión (CE), producción de leche estimada a 305 días (P305) y tiempo de transferencia del embrión (TTE), respectivamente(22).
RESULTADOS
Vacas donadoras
El número de partos (2.4 ± 0.25), los días en leche (105.3 ± 7.0) y la condición corporal (3.3 ± 0.07) fueron similares en ambas épocas de colección embrionaria (P>0.05). La producción de leche estimada a 305 días (P305) (11,609.9 ± 473.0 L) y el peso corporal (674.8 ± 18.8 kg) fueron superiores en vacas de época templada, comparados a lo observado en época cálida (9,992.7 ± 444.9 L y 612.2 ± 17.8 kg, respectivamente) (P<0.01).
Vacas receptoras
El número de partos (1.98 ± 0.09), los días en leche (86.0 ± 2.7), la condición corporal (3.3 ± 0.04) y el peso corporal (596.6 ± 10.5 kg) de las vacas receptoras el día de la transferencia embrionaria fueron similares entre épocas de transferencia (P>0.05). La P305 en vacas de época de transferencia templada (9,829.3 ± 276.8 L) fue superior (P<0.01) a la observada en vacas de época cálida (8570.2 ± 279.4 L).
Respuesta superovulatoria y colecta embrionaria
La respuesta superovulatoria, el número de cuerpos lúteos detectados al momento de la colecta embrionaria y la tasa de fertilización fueron similares en ambas épocas (P>0.05). En la época templada, el número de embriones y óvulos y la tasa de colección de embriones más óvulos por vaca fueron superiores a lo observado en la época cálida (P<0.05) (Cuadro 3). En la época cálida se observó una tendencia de mayor porcentaje de vacas a las que se les recolectó de 0 a 2 embriones y óvulos durante la época cálida (36.8 %) que en la época templada (11.8 %, P<0.07); mientras que el porcentaje de vacas a las que se les recolectó de 9 a 11 embriones y óvulos la tendencia fue mayor en la época templada (35.3 vs 10.5 %, P<0.06).
La distribución porcentual de los embriones por su estadio de desarrollo: blastocito (58.1 %); mórula (26.1 %) y retrasados (15.7 %) no fue diferente entre épocas de colección embrionaria (P>0.05). Sin embargo, el porcentaje de embriones de calidad excelente fue mayor en la época templada (62.0 %) que lo observado en la época cálida (42.0 %, P<0.01).
Tasa de gestación
La tasa de gestación de las vacas receptoras de embriones fue mayor durante la época templada (33.3 %) que durante la cálida (14.0 %; P<0.05). La tasa de gestación de las vacas receptoras de época templada fue más alta cuando recibieron embriones producidos durante la época templada (45.0 %), que en aquéllas que recibieron embriones producidos en época cálida (21.5 %, P<0.05) (Cuadro 4). Se observó una tendencia de mayor tasa de gestación de las vacas receptoras cuando recibieron embriones de calidad excelente (30.4 %), que en aquéllas que recibieron embriones de calidad buena (16.9 %; P<0.11).
No se encontró efecto de la P305 sobre la tasa de gestación de las vacas receptoras (P>0.05). La concentración sérica de progesterona de las vacas receptoras el día de la transferencia del embrión (1.93 ± 0.2 ng/ml) y el tiempo de transferencia del embrión (4.12 ± 0.15 min) fueron similares en ambas épocas (P>0.05).
DISCUSIÓN
El estrés calórico afecta la viabilidad embrionaria y el medio ambiente materno para el establecimiento de la gestación. El hecho de que la tasa de gestación de las vacas receptoras de época templada se haya reducido en más del 50 % cuando éstas habían recibido un embrión producido en época cálida, comparada con la observada en vacas que recibieron un embrión cuyo origen de producción fue de época templada, podría indicar un efecto negativo del estrés calórico sobre la viabilidad del embrión, aún cuando éste se haya desarrollado hasta el estadio de blastocisto. Este efecto adverso del estrés calórico sobre el desarrollo temprano del embrión ha sido previamente reportado en estudios in vivo(6) e in vitro(5,23,24). Otra posibilidad pudo haber sido el efecto que tiene el estrés calórico sobre el desarrollo y la calidad del folículo y el óvulo antes de la ovulación, que reduce el desarrollo y la sobrevivencia del embrión(1,3,4,5). Al respecto, se ha demostrado que la eliminación frecuente de folículos desarrollados durante el verano, permite el reclutamiento de folículos saludables e incrementa la proporción de embriones producidos in vitro hasta el estadio de blastocisto(2,3).
En el presente estudio, el efecto positivo de los embriones producidos en condiciones templadas para incrementar la tasa de gestación no fue observado en vacas receptoras en condiciones de estrés calórico; y que independientemente de la época de producción del embrión y la calidad del embrión, las vacas receptoras de época cálida presentaron una reducción de la tasa de gestación de más del 58 %, que puede deberse a la presencia de un efecto materno que impacta sobre la sobrevivencia del embrión después de la transferencia. Estos datos concuerdan con lo observado en vacas receptoras Holstein expuestas a condiciones de estrés calórico(16,23). Sin embargo, otros estudios realizados en todas las estaciones del año han informado una tasa de gestación similar en vacas Holstein(13,15) y en vacas Bos indicus(25), cuando se les transfiere embriones producidos en condiciones de termoneutralidad.
Se ha descrito que la menor fertilidad observada durante la época cálida puede ser explicada por la pérdida embrionaria temprana(23,26,27); la cual ha sido asociada cuando las vacas receptoras recibieron un embrión al inicio del verano(23); cuando se tienen alteraciones inducidas por estrés calórico en la síntesis de proteínas por el endometrio durante los primeros ocho días del ciclo estral(28,29), y con menor secreción de interferon-tau en el día 17 en conceptus en cultivo(30).
Los factores de crecimiento embriotróficos presentes en el lumen uterino(31) han sido considerados como factores de sobrevivencia para la pre-implantación de los embriones bovinos expuestos a estrés calórico. Durante el verano se han descrito menores concentraciones de insulina(32,33), del factor de crecimiento parecido a la insulina tipo 1 (IGF-1) y glucosa(33); eventos asociados al menor consumo de materia seca(34) y al balance de energía negativo(33,35). Varios estudios han observado en embriones cultivados con IGF-1 y que sufrieron un shock térmico, que estos pudieron continuar su desarrollo hasta el estadio de blastocisto(24,36,37) y mejoraron la tasa de gestación de las vacas receptoras(23). Por lo que se ha establecido que quizá con la manipulación del sistema IGF-1 con la administración de somatotropina bovina en vacas lactantes expuestas a estrés calórico se puede incrementar la sobrevivencia embrionaria, el crecimiento del concepto, la producción de interferon-tau y la tasa de preñez(38).
La producción estimada de leche a 305 días de las vacas receptoras no afectó la tasa de gestación, como ha sido informado en otros estudios(16,39); probablemente debido a la poca variabilidad de la producción de leche entre las vacas dentro de cada época de evaluación. Sin embargo, este efecto no debe de ser descartado, ya que se ha observado una relación negativa del alto nivel de producción de leche de las vacas con la pérdida embrionaria después de la transferencia de embriones(14); y con menor fertilidad observada durante el verano en hatos lecheros de Florida, EE.UU(40) y del centro – norte de México(41).
Independientemente de la época de transferencia, la calidad del embrión fue un factor que influyó sobre la tasa de gestación de las vacas receptoras, cuyo efecto ha sido documentado(16). Aún cuando el tiempo requerido para realizar la transferencia embrionaria fue similar entre épocas, se observó que un mayor tiempo de transferencia redujo la tasa de gestación; quizá debido a un mayor tiempo de contacto del embrión descongelado con el crioprotector, que probablemente cause un daño en la integridad del embrión(21).
Las concentraciones séricas de progesterona de las vacas receptoras al momento de la transferencia del embrión no tuvieron relación alguna con la tasa de gestación, lo que concuerda con lo informado en otros estudios(10,39), por lo que esta variable tiene un uso práctico limitado en programas de transferencia embrionaria.
Un menor número de embriones y óvulos recolectados por vaca durante la época cálida, aún cuando el número de cuerpos lúteos no fue comprometido, podría relacionarse con fallas en la ovulación, cuyo evento se ha observado con mayor frecuencia en vacas que estuvieron expuestas a estrés calórico(16,33,42) y en aquéllas cuyas concentraciones de insulina e IGF-1 estuvieron bajas(43).
Otra posible causa de la menor colecta embrionaria en vacas de época cálida podría ser que los cuerpos lúteos detectados al momento de la colección embrionaria hubieran sido en realidad folículos luteinizados, como fue observado en vacas superovuladas con hipotiroidismo inducido(44).
El estrés calórico no fue determinante para afectar la tasa de fertilización, cuyo dato coincide con lo descrito, tanto en estudios in vivo realizados en vaquillas superovuladas que fueron expuestas a altas temperaturas durante 10 h al inicio del estro(12); como en estudios de fertilización in vitro, al utilizarse óvulos obtenidos de vacas expuestas a estrés calórico durante el verano(4).
La ausencia de efecto del estrés calórico sobre el desarrollo embrionario fue similar a lo informado en vacas superovuladas con altos niveles de producción de leche durante el verano(12). En contraste, en estudios in vitro se observó un menor porcentaje de embriones que alcanzaron el estadio de blastocisto cuando estos fueron expuestos a temperaturas fluctuantes en los primeros ocho días después de la fertilización, similares a las observadas en las vacas en un día de verano(45). Por lo que quizá el nivel de estrés calórico del presente estudio no fue suficiente para comprometer el desarrollo embrionario, pero si para afectar la calidad embrionaria, como ha sido descrito por otros autores(16).
CONCLUSIONES E IMPLICACIONES
El hecho que los embriones producidos durante la época templada haya incrementado la tasa de gestación de las vacas receptoras en condiciones de confort térmico, y que dicho evento no haya sido observado en vacas que recibieron embriones producidos durante la época cálida, implica quizá un daño en la calidad del óvulo y en el desarrollo embrionario por efecto del estrés calórico, que pudo haber afectado la viabilidad del embrión para asegurar la gestación de la vaca receptora. La menor tasa de gestación observada en vacas receptoras de épocas cálidas, independientemente de la época de producción del embrión y su calidad, puede implicar un efecto materno asociado al estrés calórico sobre la viabilidad del embrión después de la transferencia. Es por esto que quizá el estrés calórico pudiera ser estratégicamente manejado para disminuir su efecto detrimental sobre la tasa de gestación de las vacas receptoras, especialmente alrededor del tiempo de la transferencia del embrión.
AGRADECIMIENTOS
La investigación fue apoyada por el CONACyT (31457-B); Fundación Produce Aguascalientes A.C. y la Asociación de Productores de Leche del Estado de Aguascalientes. Se agradece en forma especial a los propietarios y personal de campo de las explotaciones lecheras de “Granja El Sol” y “Rancho Las Palomas”, quienes colaboraron con la aportación de los animales y facilitaron las labores de manejo del ganado.






")










")



